
Proboscidea is
the order of mammals with trunks, and their more primitive relatives
with “proto-trunks”. Today the only living members of this order
are in the family Elephantidae, the
living Asian, African savannah and forest elephants. These are
magnificent megafauna in themselves, treasures of any zoo which has
them, butchered for their incisors and occasionally used in human
warfare. But there is much more to Proboscidea than
the living elephants as this order has a total of around 164 mostly
extinct species, which diversified into all sorts of bizarre forms,
such as the four tusked deinotheres and the shovel-tusked
Platybelodon. These
fantastical, mighty beasts are only known to us through the fossil
record. It is tempting to compare the largest Proboscideans, to the
herbivorous dinosaurs, especially the sauropods, as they fill the
niche for large browsing animal. I have no chance of discussing all
these species here, but I will mention some of the most outlandish
and interesting Proboscideans less familiar to us than elephants and
mammoths.
Animals
with 3m long teeth don't drop out of the sky, they as an order have a
history stretching back into the Paleocene, starting with an
apparently humble creature the size of a rabbit. The evolutionary
process basically involves a gradual increase in body size, growth of
tusks and size of head. The heavy tusked head could not be supported
by a long neck, and as their legs were long they could not reach the
ground with their mouth to eat and drink. This problem was “solved”
by the elongation of the muscular nasal cavity, forming a trunk.
These processes were not so smooth and continuous, and the journey
had many turns from this path.

A reconstruction of Eritherium.
One
of the earliest Proboscidean is Eritherium
which lived 60 million years ago, the remains of this creature
was first found in phosphate deposits in Morocco in 2012. The
Eritherium remains show no sign of a trunk, which is formed by
the evolutionary fusion of upper lip and nostril, which would be
evident in an enlarged naval cavity. It is a Proboscidean without a
proboscis, but probably had a very mobile upper lip, a little like
the unrelated tapirs. So what makes this animal a Proboscidean? A
number of features which were elaborated in later Proboscideans are
present in Eritherium, including enlarged incisors, which
would eventually sprout forth to create tusks, and simple lophodont
molars (molars with ridges perpendicular to the jaw line). It was 5kg
in weight, but this outweighs most other Paleocene mammals, many of
which were decided shrew-like. Eritherium was probably
somewhat aquatic, like many of the Proboscideans including the swamp
dwelling American Mastodon and modern elephants who use their trunks
as “snorkels” in order to swim up to 48km offshore. It
is likely that the common ancestor of Proboscideans, Sirenians (sea
cows) and the extinct Desmostylia were fully aquatic.

A reconstruction of Phiomia
A
later group of Proboscideans is Phiomia, a
direct relative of modern elephants, which lived between 35
and 25 million years ago. This was a larger animal, 2m high at the
shoulder. Remains show evidence of a short rudimentary trunk (based
on the larger nasal cavity) and enlarged incisors on the lower and
upper jaws, meaning it had two pairs of tusks. Tusks are simply
enlarged incisors, about a third of it's length, the pulp cavity, is
embedded in the skull. The visible tusk is the ivory made of dentine
covered by enamel. After it has shed it's milk tusks a Proboscidean
maintains and grows it's tusks throughout it's life. In observed
species the male has the longest tusks, as befitting their primary
use in fighting, but they are and were also used to strip bark,
defend from predators and in species like the woolly mammoth they may
have been used to break up ice to find food.

A skull of Deinotherium at the Natural History Museum
The
branches of evolutionary tree which did not lead to elephants contain
more baroque Proboscideans, including two groups which lived at
approximately the same time, Deinotherium and
Platybelodon. The skull of Deinotherium is
a disturbing thing to bump into in a museum, no doubt horrifying for
an ancient person, with no ideas about deep time, to find in the
wild. The skull appears to have two horns curved backwards out of
it's chin, like a demonic beard presumably used for gouging. If an
ancient person who expects all animals to have forward facing eyes
like they do, sees the enlarged nasal cavity at the front of the
skull, it could be mistaken for the skull of a monster. Indeed, the
ancient myths of the Cyclops could be inspired by the discovery of
Proboscidean fossils in Greece. Deinotherium itself lost it's
upper pair of tusks and maintained the lower tusks (the opposite of
modern elephants). As you can tell from a reconstruction of these
creatures, the lower tusks hid quite discretely under the trunk which
filled the nasal cavity at the front of the animals head, making it
look less horrifying. They were quite primitive, lacking the
sequential teeth eruption of later Proboscideans.
The
skull of Deinotherium is a disturbing thing to
bump into in a museum and no doubt horrifying for an ancient person,
with no ideas about deep time, to find in the wild. The skull appears
to have two horns curved backwards from it chin, like a monstrous
beard, used for gouging. Indeed, the ancient myths of the Cyclops
could be inspired by the discovery of Proboscidean fossils in Greece,
due to the fused external naris resembling an eye socket. Even the
name Deinotherium is from the Greek for “terrible beast”.
In reality, Deinotherium lost it's upper tusks and maintained
it's lower tusks (the opposite of modern elephants), and probably
used these curved tusks to scrap bark from tree. The lower tusks
would have hid quite discretely under the trunk which extended from
the external naris, making it look less horrifying.
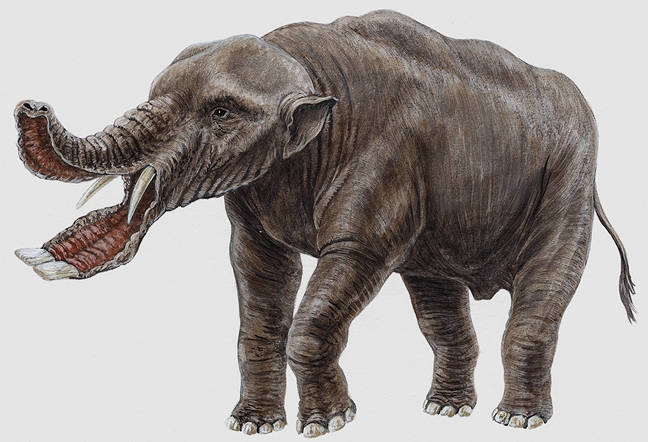
A reconstruction of Platybelodon
Platybelodon
is in the same family as Gomphotherium and is
not a direct ancestor of elephants. It lived about 20 – 8 million
years ago and retained all four tusks. The lower tusks
flattened out so the each tusk met and formed a sort of “shovel”
shape with a deep scoop at the end, which only developed in
adulthood. There is speculation about the use of it's tusks. It was
originally thought that it they used their “shovel” to scoop
through the mud to collect plants; a semi-aquatic lifestyle familiar
to Proboscideans. The two lower tusks end with a V-shaped sharpened
tip, analysis of the pattern of wear suggests they were used in a
scythe-like manner to cut down branches and to strip bark from trees.
Palaeontologists removed them from their presumed habitat of
lake-side bogs and place them in a more arboreal habitat. The shovel
tusks were an adaptation to a crowding niche, as with several genera
of Proboscidean in this area at the time, Platybeldon had to
specialise to survive.

A reconstruction of Gomphotherium
Another
key part of the future elephant physiology was put in place in
another of the elephants' direct ancestors, Gomphotherium,
living from 20 to 15 million years ago. They retained the four
tusks, with the lower tusks being slightly flattened, and show a
basic form of sequential tooth development. This is when, due to the
stresses caused by eating grasses containing silicon particles, a
tooth erupts from behind the existing teeth to replace the one which
had eroded at the front. The erupted tooth would be larger than the
previous ones, therefore allowing the jaw to continue to grow
throughout the animals life. Gomphotherium probably had three
teeth in each side of it's jaw, but later species had only one tooth
in each side of the jaw. Gomphotherium was a member of the
well travelled family Gomphothere, which includes the genera
Cuvieronius and Stegomastodon, some species of which
travelled as far as South America.

A sketch of American Mastodon Molars, my work
Mastodons
were, despite their common confusion with mammoths, very different
animals, Mastodons diverged from the line that would lead to
elephants after Phiomia, separating them from mammoths by
about 20 million years of evolution. The earliest Mastodons were the
Losodokodon which lived from 27 to 24 million years ago in
East Africa, later radiating throughout Europe, Asia and North
America. The most famous Mastodon species is the American Mastodon,
whose genus Mammut arrived in North America 11 million years
ago. The American Mastodon was around 2.7 metres at the shoulder,
small compared to the neighbouring Columbian Mammoth. It was quite
stocky with a deep chest and probably quite muscular. It's tusks were
up to 2.5 metres long in adult males and curved upwards and slightly
outwards, less elaborate than tusks of mammoth. The name Mastodon
means “breast tooth”, showing the lumpy nature of their molars.
Elephant and mammoth molars are relatively flat and ridged, whereas
mastodon molars have rounded cusps, which caused early naturalists to
speculate that they were terrifying beasts who caught prey with their
tusks. The reason each of these groups have significantly different
teeth is because they were used to eat different foods, none of these
animal foods. Mastodon teeth were used to crush leaves and twigs in
forests, whereas mammoths grazed grassland.

My drawing of a cave painting of a woolly mammoth
The
genus believed to be the ancestors of modern elephants and mammoths
is Primelephas. These creatures had four tusks,
though these were smaller than in Gomphotherium. This
is because Primelephus did not need them to shovel though the
mud, it had moved to a grassland habitat similar to modern elephants
on the savannah or mammoths on the steppe. This genus split into
three genera, Loxodonta (African elephants), Elephus (Asian
elephants) and Mammuthus (mammoths). Mammuthus belongs
to the Elephantidae family, making it as much of an elephant
as Loxodonta and Elephus.
Bibliography
Understanding
proboscidean evolution: a formidable task- Jeheskel
Shoshani www.sciencedirect.com/science/article/pii/S0169534798014918
I
found the exhibition Mammoths: Ice Age Giants at the Natural History
Museum very informative, as well as the book of the same name by
Adrian Lister.
This was an entry to Rockwatch's Young Writer 2014 competition.





























{kind=link}